Щитовидная железа имеет вид бабочки, крылья которой представлены левой и правой долями, соединенными между собой перешейком. Размеры каждой доли составляют 2,5-4 см в длину, 1,5-2 см в ширину и 1-1,5 см – толщины. В 20-25% выявляется пирамидальная доля, располагающаяся над перешейком и представляющая собой небольшое количество тироидной ткани. Реже выявляются добавочные доли щитовидной железы, располагающиеся в треугольнике, основанием которого является подъязычная кость, а вершиной – щитовидный хрящ. Щитовидная железа заключена в соединительнотканную оболочку, состоящую из наружной и внутренней капсулы, в пространстве между которыми находятся сосуды, возвратный нерв и околощитовидные железы.
Синтез тироидных гормонов осуществляется в фолликулах, которые представляют собой функциональную и морфологическую единицу щитовидной железы. Форма и размеры фолликулов зависят от функционального состояния железы, их диаметр колеблется от 15 до 500 мкм. Стенки фолликулов состоят из одного слоя эпителиальных клеток (тироцитов), верхушки которых направлены в просвет фолликула, а основания прилежат к базальной мембране. Апикальная часть тироцита имеет микроворсинки, образующие псевдоподии. Мембрана этой части тироцита содержит ферменты – пероксидазу, аминопептидазу и обладает способностью генерировать перекись водорода, необходимую для биосинтеза тироидных гормонов. Базальная часть мембраны тироцита содержит Na+, K+-АТФазы и рецепторы к ТТГ. Каждый фолликул окружен сетью лимфатических сосудов и капилляров, которые являются продолжением в основном верхней и нижней артерий щитовидной железы. Скорость кровотока в щитовидной железе составляет 5 мл/г•мин. Волокна симпатической части вегетативной нервной системы (далее – симпатическая нервная система), сопровождающие сосуды, оканчиваются на стенке как капилляров, так и фолликулов. Щитовидная железа получает иннервацию и от парасимпатической части вегетативной нервной системы (далее – парасимпатическая нервная система). Фолликулы (20-40) образуют дольки, отделенные друг от друга соединительной тканью.
Помимо фолликулярных клеток, в щитовидной железе имеются так называемые С-клетки, или парафолликулярные клетки, секретирующие кальцитонин – один из гормонов, регулирующих гомеостаз кальция. Эти клетки выявляются в стенке фолликулов или в интерфолликулярных пространствах.
Полость каждого фолликула заполнена коллоидом, состоящим в основном из тироглобулина – гликопротеида (мол. м. 660 кДа). Молекула тироглобулина включает 5496 аминокислотных остатков и содержит 140 тирозиновых остатков и около 10% углеводов (галактоза, манноза, фукоза, сиаловые кислоты, N-ацетилглюкозамин, хондроитин сульфат). Содержание сиаловых кислот в тироглобулине непостоянно и увеличивается с повышением содержания йода. Показано, что при папиллярном раке щитовидной железы содержание сиаловых кислот значительно снижено (J. Sinadinjvic и соавт. 1992).
Синтез тироглобулина контролируется геном, локализованным на длинном плече 8-й хромосомы и являющимся наибольшим из известных генов человека, состоящим из более чем 300 kb, причем лишь 64 kb приходится на 42 экзоны и интроны гена. ТТГ стимулирует транскрипцию гена тироглобулина, тогда как гипофизэктомия или введение экзогенных тироидных гормонов ингибируют транскрипцию указанного гена. Тироглобулин синтезируется на рибосомах шероховатой эндоплазматической сети тироцита в виде претироглобулина, далее переносится в цистерны, где формируются вторичные и третичные структуры тироглобулина, а также углеводные компоненты молекулы, которые, как указано выше, представлены здесь в основном моносахаридами. Из цистерн шероховатой эндоплазматической сети тироглобулин транспортируется в комплекс Гольджи, где происходит окончательное образование углеводных компонентов. Далее тироглобулин перемещается к апикальной части клетки, образуя пузырьки. Путем экзоцитоза содержимое пузырьков освобождается в просвет фолликула. Здесь на апикально-коллоидном барьере под влиянием ферментов мембраны апикальной части тироцита происходит йодирование тироглобулина, с последующим его “складированием про запас” в форме коллоида. Из всех тирозиновых остатков, входящих в молекулу тироглобулина, лишь 1/5 подвергается йодированию и лишь 1/6-1/8 последних превращаются в дальнейшем в йодтиронины. В нормальных условиях транспорт тироглобулина совершается не только в просвет фолликула, но и в обратном направлении. Тироглобулин, обнаруживаемый в сыворотке крови, является гетерогенным вследствие сплайсинга мРНК (F. Bertaux и соавт., 1991) и его секреция в кровяное русло стимулируется ТТГ, где период полураспада составляет от 3 до 28 ч.
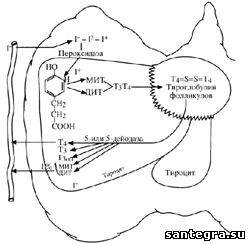
Процесс биосинтеза тироидных гормонов (схема 21) можно разделить на 4 стадии.
|
|
Схема 21. Синтез и секреция тироидных гормонов. Объяснение в тексте.
Включение йода в щитовидную железу. Йод в виде органических и неорганических соединений поступает в желудочно-кишечный тракт с пищей и питьевой водой. Йод всасывается в кишечнике в виде йодидов. Установлено, что суточное потребление йода составляет 150-200 мкг, в странах Европы до 300 мкг, а в США – до 500 мкг. |
Йодиды с током крови достигают щитовидной железы, ткань которой обладает уникальной способностью захватывать и концентрировать йодид со скоростью около 2 мкг/ч. Если поступление йода в организм снижается ниже 100 мкг/сут, развивается вначале гиперплазия, а затем нетоксический зоб щитовидной железы.
Электронномикроскопические исследования показали, что уже через 30-40 с после введения 125-I отмечается его накопление по периферии просвета фолликула в непосредственной близости от апикальной части клетки. В желудочно-кишечный тракт йодиды секретируются слюнными железами и слизистой оболочкой желудка, где содержание йодидов в 20-30 раз выше, чем в плазме. Однако в этих органах, как и в молочной железе, отсутствуют ферменты, необходимые для органификации йода.
Клеточные мембраны тироцитов, осуществляющие захват йодидов (I-), не могут отличать моновалентные анионы один от другого и поэтому способны захватывать наряду с йодидом и другие анионы, несущие отрицательный заряд: SCN-, CLO-4, TcO-4. Если в организм избыточно поступают эти анионы, происходит их накопление в щитовидной железе и путем конкуренции угнетается поглощение йода. В таких случаях недостаточный захват йодидов щитовидной железой приводит к снижению их количества в этом органе и как следствие – к недостаточному синтезу тироидных гормонов.
Транспорт йодида через мембрану тироцита является активным, требующим затрат энергии процессом, при котором йодид поступает из среды с меньшей концентрацией (плазма крови) в среду с высокой концентрацией (ткань щитовидной железы). Концентрация свободного йода в щитовидной железе в 30-40 раз выше, чем в плазме крови.
Транспорт йода через мембрану клетки осуществляется с участием Na+-, K+-АТФазы. Установлено, что некоторые сердечные глюкозиды, угнетающие активность АТФазы в щитовидной и подчелюстных железах, угнетают также транспорт йодида в щитовидную железу. Ионы натрия (Na+) тоже влияют на транспорт йодида, активируя процессы выхода йодида из клеток щитовидной железы. Одним из возможных переносчиков йодида через клеточную мембрану могут быть фосфолипиды.
Не исключено, что перенос йодида через мембрану тироцита осуществляется специфическим, еще не идентифицированным белком. Это предположение подкрепляется тем, что процесс захвата и транспорта йодида находится под генетическим контролем, а также тем, что повышение поглощения йода щитовидной железой под влиянием ТТГ происходит лишь через несолько часов, а не сразу после воздействия гормона.
Встречаются клинические варианты гипотироза, обусловленные недостаточностью образования тироидных гормонов вследствие дефекта в системе, осуществляющей захват йодида из плазмы крови и транспорт его через мембрану тироцита. В таких случаях у обследованных выявляется низкое поглощение введенного в организм радиоактивного йода щитовидной железой, а также низкое содержание его в слюне, желудочном соке или молоке кормящих матерей. В этой связи необходимо отметить, что наряду со щитовидной железой, слюнными железами и слизистой желудка способностью концентрировать йод обладают также и молочные железы, которые могут накапливать йод в 10 раз больше и более активно по сравнению со щитовидной железой.
Органификация (окисление) йода или йодирование тирозина в тироглобулине. Cледующим этапом после захвата йода щитовидной железой является синтез тиродных гормонов, котрый начинается с быстрой фиксации йода в молекулу тирозина. Однако, прежде чем поступивший в щитовидную железу йодид будет использован для синтеза тироидных гормонов, он должен быть окислен до активной формы при помощи фермента тиропероксидазы и перекиси водорода (H2О2). Перекись водорода генерируется дигидроникотинамид аденин динуклеотид фосфатом (NaDPH), который при участии соответствующей оксидазы окисляется в присутствии ионов кальция и этот процесс стимулируется ТТГ. Тиропероксидаза является микросомальным антигеном, к которому образуются антитела при аутоиммунных заболеваниях щитовидной железы и, в частности, при аутоиммунном тироидите. Тиропероксидаза является мембраносвязанным гемопротеином, который очень нестабилен и поэтому трудно поддается очистке. Установлено, что молекула тиропероксидазы состоит из 933 аминокислот и ее гликозилированная форма имеет молекулярную массу 107 кДа. Активированный таким образом йодид (I+) способен йодировать молекулу тирозина с образованием монойодтирозина (МИТ) или дийодтирозина (ДИТ). При помощи этой тиропероксидазной системы щитовидная железа использует каждый поступающий в нее атом йода и препятствует возможному возвращению йодида в кровяное русло. Однако тиропероксидаза может катализировать йодирование тирозина, входящего в состав других белков (альбумин и др.). Такой метаболически неактивный белок может выделяться в кровообращение, что приводит к большой “утечке” йода из щитовидной железы.
Процесс образования МИТ и ДИТ происходит на боковых цепях молекулы тироглобулина, несущих тирозиновые остатки. Йодирование тироглобулина осуществляется на границе между апикальной частью тироцита и коллоидом, содержащимся в просвете фолликула
Процесс конденсации. В заключительной стадии гормоносинтеза МИТ и ДИТ под влиянием окислительных ферментов конденсируются с образованием биологически активных гормонов щитовидной железы: трийодтиронина-Т3 и тироксина-Т4. При конденсации двух молекул ДИТ образуется Т4; если происходит конденсирование между молекулами МИТ и ДИТ, образуется Т3. В настоящее время получены убедительные данные, позволяющие считать, что процесс конденсации осуществляется в 3 этапа под влиянием тиропероксидазы. Сначала йодтирозины окисляются с помощью этого фермента в активную форму. Активированные таким образом йодтирозиновые остатки внутри той же молекулы тироглобулина превращаются в интермедиатные квинольные эфиры. Последние через конверсию боковой цепи аланина до дегидроаланина образуют тиронины. На этой заключительной стадии необходима димерная структура тироглобулина, позволяющая осуществлять процесс конденсации внутри молекулы тироглобулина.
Таким образом, тиропероксидаза контролирует все этапы биосинтеза тироидных гормонов. Однако ДИТ непосредственно может стимулировать процесс конденсации, но механизм этого явления пока неясен. Хотя и редко, но встречаются случаи гипотироза, когда процесс биосинтеза тироидных гормонов протекает нормально лишь до стадии конденсации, в то время как процесс образования Т3 и Т4 резко заторможен. Образование и конденсация МИТ и ДИТ происходят на мембране в апикальной части тироцита, где локализуется тиропероксидаза. По мере того, как на молекуле тироглобулина все остатки тирозина йодируются с последующим образованием тирозинов и тиронинов, она перемещается в просвет фолликула, где и происходит их накапливание “про запас”. Большая часть фолликулов щитовидной железы предназначена для хранения тироглобулина и количество в нем тироидных гормонов таково, что если полностью блокировать биосинтез тироидных гормонов, то запасов Т3 и Т4 будет вполне достаточно для поддержания эутироидного состояния в течение месяца. Считается, что тиростатические препараты ингибируют тиропероксидазу и блокируют образование тироидных гормонов не только на стадии образования МИТ и ДИТ, но даже в большей степени на стадии конденсации и образования Т3 и Т4.
Высвобождение (секреция) гормонов щитовидной железы. При снижении уровня тироидных гормонов в сыворотке крови “срабатывают” центры, контролирующие секрецию ТТГ и приводящие к стимуляции его высвобождения. ТТГ, комплексируясь с рецепторами тироцитов, активирует аденилатциклазу и увеличивает образование цАМФ. Действие цАМФ на высвобождение тироидных гормонов осуществляется через активирование (или синтез) ферментов (протеазы, эндопептидазы, гликозидгидролиазы, фосфатазы и др.), контролирующих протеолиз, и путем непосредственного влияния на процессы транспорта тироглобулина из просвета фолликула к лизосомам клетки, осуществляющим протеолиз тироглобулина. Уже через 10 мин после действия ТТГ в апикальной части тироцитов образуются псевдоподии, которые путем эндоцитоза захватывают капельку коллоида. Почти одновременно с началом эндоцитоза коллоида также под влиянием ТТГ плотные гранулы, по существу являющиеся лизосомами, мигрируют от базальной мембраны к апикальной части клетки и сливаются с капельками коллоида, образуя фаголизосомы, которые содержат эстеразу и фосфатазу, но преимущественно катептические протеазы. В результате гидролиза тироглобулина высвобождаются Т3 и Т4, свободные аминокислоты, которые поступают в периваскулярное пространство, а лизосомальные белки остаются в клетке.
В процессе гидролиза тироглобулина наряду с Т3 и Т4 происходит высвобождение МИТ и ДИТ, которые подвергаются дейодированию под действием йодтирозиндейодазы, которая выявляется в митохондриях и микросомах и является NADPH-зависимым флавопротеином. Высвободившийся таким образом йодид вновь используется щитовидной железой в биосинтезе тироидных гормонов. В случае снижения активности фермента йодтирозиндейодазы не происходит дейодирования МИТ и ДИТ и они в избыточном количестве поступают в кровь и выделяются с мочой. Вследствие избыточной потери йода с мочой (в форме МИТ и ДИТ) появляется отрицательный баланс йода в организме, что приводит к недостатку йода, необходимого для биосинтеза тироидных гормонов и развитию гипотироза.
Протеолиз тироглобулина ингибируется избытом йода и литием. Небольшое количество тироглобулина не подвергается гидролизу и высвобождается из тироцитов одновременно с гормонами щитовидной железы. Однако при некоторых заболеваниях (подострый тироидит, тиротоксикоз, зоб и особенно злокачественные опухоли щитовидной железы) высвобождение тироглобулина в кровь повышается. Концентрация тироглобулина в сыворотке крови составляет 16 мкг/л, но у больных с алкогольным циррозом его уровень повышен более чем в 2 раза. Однако при остром гепатите концентрация тироглобулина в крови в пределах нормы.
Биосинтез тироидных гормонов полностью находится под контролем ТТГ. Установлено, что взаимодействие ТТГ с соответствующим рецептором активирует аденилатциклазу и накопление цАМФ. В активировании аденилатциклазы принимает участие также специфический белок Gs, т.е. ГТФ-связывающий белок. В щитовидной железе человека также активируется фосфолипаза С и гидролиз фосфатидил инозитол 4,5-бифосфата. Если у животных (собаки) ТТГ стимулирует все этапы биосинтеза тироидных гормонов (захват и транспорт йода, синтез тироглобулина, органификацию йода, синтез и высвобождение тироидных гормонов), то у человека в процессах органификации и синтеза тироидных гормонов, помимо ТТГ, участвует фосфолипаза С и Са++-ионизированный кальций. цАМФ участвует также в процессах стимуляции экспрессии генов, контролирующих синтез белка, осуществляющего транспорт йода, а также генерацию перекиси водорода, тироидную пероксидазу и тироглобулин (E. Raspe и J.E. Dumont, 1995). Кроме того, действие ТТГ и цАМФ на пролиферацию и функцию тироцитов опосредуется при обязательном присутствии ИФР-1 (пермиссивное действие)
Сравнительно недавно было показано, что йодная недостаточность сама по себе приводит к развитию зоба и нарушению функции щитовидной железы, причем выраженность этих нарушений находится в прямой зависимости от степени йодной недостаточности в организме. При этом снижается количество интратироидального йода и увеличивается отношение МИТ к ДИТ, Т3 к Т4. Повышается уровень ТТГ в сыворотке крови. Наряду с увеличением щитовидной железы появляются симптомы гипотироза. В случае наличия йодной недостаточности с момента рождения имеет место выраженная степень гипотироза, вплоть до кретинизма.
Поступившие из щитовидной железы в кровь Т3 и Т4 связываются с белками крови (тироксинсвязыающий глобулин, транстиретин или тироксинсвязывающий преальбумин и альбумин), осуществляющими транспортную функцию.
Тироксинсвязывающий глобулин синтезируется в печени и имеет мол. м. 57 кДа. Он является главным белком, связывающим тироидные гормоны в крови. Показано, что более 70% всех тироидных гормонов транспортируются в связанном с этим белком состоянии. Молекула тироксинсвязывающего глобулина включает 4 углеводных цепи и около 10 сиаловых кислот. Она имеет одно связывающее место для Т4 или Т3. Тироксинсвязывающий глобулин связывает и транспортирует 75% тироксина и 85% трийодтиронина. Период его полураспада 5 дней и скорость разрушения 15 мг/сут. Концентрация тироксинсвязывающего глобулина в крови составляет 16-29 мкг/мл или 286-552 нмоль/л. Тироксинсвязывающий глобулин более прочно связывает тироксин, в то время как связь с трийодтиронином в 4-5 раз слабее по сравнению с Т4.
Транстиретин (тироксинсвязывающий преальбумин) имеет мол. м. 55 кДа и относится к глобулярным полипептидам. Его период полураспада составляет 2 дня и скорость разрушения 650 мг/сут. Концентрация транстиретина в крови составляет 120-240 мг/л или 2250-4300 нмоль/л. Этот белок связывает около 10-15% Т4 и менее 5% Т3. Прочность связи транстиретина с тироксином значительно уступает связи тироксинсвязывающего глобулина с Т4. Связь же транстиретина с Т3 очень слабая и его способность связывать Т3 в 10 раз ниже по сравнению с Т4.
Альбумин имеет период полураспада 15 дней, скорость разрушения 7000 мг/сут. В связи с высокой концентрацией в сыворотке крови (3500 мг/100мл) альбумин обладает большой связывающей способностью и связывает около 10% Т4 и несколько меньше Т3. Его связь с тироидными гормонами наименее прочная из всех трех белков-транспортеров тироидных гормонов. Поэтому он является основным источником свободных тироидных гормонов на периферии. При циррозе печени, который сопровождается гипоальбуминурией, наблюдается сниженное содержание Т3 и Т4 в сыворотке крови, но при этом свободная фракция Т3 и Т4 в пределах нормы и у больных остутствуют клинические проявления гипотироза.
Таким образом, большая часть тироидных гормонов, а именно 99,96% Т4 и 99,6% Т3 находится в связанной с белками крови форме. Свободная фракция составляет лишь 0,04% для Т4 и 0,4% для Т3 и именно она обусловливает биологическое действие тироидных гормонов.
Необходимо отметить, что встречаются наследственные заболевания, при которых нарушается синтез транспортных белков. Их изменение выражается либо увеличением количества белков в 3-5 раз, либо их снижением вплоть до полного отсутствия, либо возникновением чрезмерно прочной связи между тироксином и белком. Врожденная недостаточность тироксинсвязывающего глобулина встречается у 1:2500-3000 новорожденных в виде 2 форм: почти у 40% коренных жителей Австралии; у африканских пигмеев, жителей Микронезии и Индонезии. Обе формы передаются Х-связанным путем. Характерным для этих состояний является низкое содержание Т3 и Т4 в сыворотке крови, но при этом отсутствуют клинические признаки гипотироза, так как концентрация свободного Т3 и Т4 в пределах нормы. Описаны больные с высоким содержанием Т4 и Т3 вследствие повышенного образования тироксинсвязывающего глобулина. Однако при нормальном уровне свободного Т3, Т4 и ТТГ у них имеет место эутироидное состояние. Описаны также семейные наблюдения, при которых повышенный уровень общего Т4 сочетается с высоким содержанием транстиретина в крови и нормальной концентрацией свободного Т3 как результат повышенного содержания транстиретина. При этом указанная патология у некоторых членов семьи сочетается с глюкагономой или другой опухолью островка поджелудочной железы. И, наконец, описана семейная дисальбуминемическая гипертироксинемия, наследуемая аутосомно-доминантным путем, проявляющаяся высоким уровнем общего Т4, при нормальной концентрации свободного Т4 в сыворотке крови. При этой патологии альбумин имеет селективную повышенную аффинность лишь к Т4, тогда как связывание его с Т3 в пределах нормы. Клинически такие больные находятся в эутироидном состоянии.
Роль белков, связывающих тироидные гормоны, в организме велика. Они связывают избыточное количество этих гормонов, ограничивая в строгих пределах фракцию свободных гормонов, и тем самым, с одной стороны, предупреждают потерю их через выделительную систему (печень и почки), а с другой стороны- регулируют скорость доставки тироидных гормонов на периферию, где они оказывают основное метаболическое действие.
Чем вызвано наличие нескольких видов тироксинсвязывающих белков? Тироксинсвязывающий глобулин, как указано выше, связывает наиболее прочно по сравнению с другими белками как Т4, так и Т3 и является стабильным, относительно инертным резервуаром тироидных гормонов. Транстиретин и альбумин представляют собой как бы лабильную фракцию тироидных гормонов, способную поставить при различных стрессовых ситуациях необходимое количество свободных тироидных гормонов. Различная степень связывания Т3 и Т4 с тироксинсвязывающими белками объясняет, хотя бы частично, более быстрое периферическое действие Т3.
Концентрация тироксинсвязывающих белков может изменяться под влиянием различных факторов. Так, повышению содержания этих белков способствуют прием эстрогенов, беременность, гипотироз, острый инфекционный гепатит. Снижение тироксинсвязывающего глобулина наблюдается при лечении глюкокортикоидами, андрогенами, а также при циррозе печени, нефротическом синдроме, тиротоксикозе и недостаточности белкового питания.
Наибольшее биологическое значение имеет Т3, который активнее Т4 в 4-5 раз. В течение длительного времени считалось, что Т4 и Т3 в равной степени принимают участие в обмене веществ. Однако было показано, что на периферии Т4 конвертируется (переходит) в Т3 и биологическое действие тироидных гормонов более чем на 90-92% осуществляется за счет Т3. Использование радиоиммунологического метода для определения концентрации Т3 в сыворотке крови позволило установить, что около 80% циркулирующего Т3 в сыворотке крови является производным от Т4 вследствие его периферического монодейодирования и только 20% Т3 сыворотки непосредственно образуется в щитовидной железе.
Таким образом, основная роль тироксина в организме заключается в том, что он является своего рода источником или, правильнее, прогормоном Т3. В процессе монодейодирования Т4 конвертируется как в Т3, так и в обратный (реверсивный) трийодтиронин, являющийся биологически неактивным гормоном. В норме в течение суток щитовидная железа секретирует около 100 нмоль Т4; около 5 нмоль Т3 и меньше чем 5 нмоль обратного Т3 (обТ3).
Периферическое монодейодирование Т4 осуществляется с помощью ферментов – дейодаз (дейодиназ) и в настоящее время идентифицировано 3 типа таких дейодаз: тип I, 5’-дейодаза; II тип, 5’-дейодаза, III тип, 5’-дейодаза.
Тип I 5’-дейодаза наиболее широко представлена в тканях (печень, почки, скелетные мышцы и мышца сердца, щитовидная железа и др.). Она содержит селеноцистеин, который обусловливает активное место дейодирования. Эта дейодаза обеспечивает прохождение Т3 в плазму и, видимо, этим объясняется повышение ее уровня при тиротоксикозе и снижение при гипотирозе. Интересно, что пропилтиоурацил ингибирует активность этого фермента, тогда как другие тиростатики (мерказолил, метимазол) оставляют его интактным. Этим фактором объясняется более быстрый терапевтический эффект пропилтиоурацила при тиротоксикозе по сравнению с действием мерказолила.
Тип II 5-дейодаза представлена в ЦНС и гипофизе. Она поддерживает постоянную концентрацию внутриклеточного Т3, необходимого для нормального функционирования нервной и эндокринной систем. Этот фермент нечувствителен к пропилтиоурацилу, но высоко чувствителен к циркулирующему уровню Т4. Между активностью фермента и содержанием Т4 имеется обратная зависимость, которая представляет собой “защитный” механизм нервной системы от избыточного образования Т3 в тех случаях, когда уровень Т4 повышается (т.е. при увеличении концентрации Т4 активность типа II 5’-дейодазы снижается).
Тип III 5’-дейодаза выявляется в плаценте и глиальных клетках нервной системы. Она конвертирует Т4 в обт 3. Активность этого фермента повышена при тиротоксикозе и снижена при гипотирозе.
При оценке содержания тироидных гормонов в крови следует учитывать влияние ряда экзогенных и эндогенных факторов на конверсию Т4 в Т3. Снижение конверсии Т4 в Т3 наблюдается при низкокалорийной диете, заболеваниях печени, системных заболеваниях, недостаточности содержания в продуктах питания селения (недостаточность селения), а также при приеме пропилтиоурацила, анаприлина, глюкокортикоидов, амиодарона и различных рентгеноконтрастных веществ, содержащих йод (иподат натрия, йопоновая кислота и др.).
Основное количество (более 80%) циркулирующего в кровообращении Т4 конвертируется с помощью дейодаз в Т3 (35%) или в обТ3 (45%). К настоящему времени неизвестны факторы, лимитирующие и определяющие в каждом конкретном случае конверсию Т4 в Т3 или в обТ3, но ясно, что эти механизмы связаны со способностью “точно определять” свободный уровень тироидных гормонов в цитозоли клетки и в крови, состояние метаболизма и связанную с этим скорость термогенеза. Интегрированная информация о состоянии перечисленных показателей поступает на “распределительный щит” и изменяет активность перечисленных дейодаз. При последующем дейодировании как Т3, так и обТ3 образуются другие тиронины, обладающие значительно более низкой биологической активностью. Так, если биологическую активность l-Т4 принять за 100%, то у Т3 – 300-800%; обТ3-меньше 1%; 3,3’-Т2 (3,3’-дийодтиронин) – 1-3%; 3,5-Т2 (3,5-дийодтиронин) – 7-11%; 3’, 5’-Т2 (3’, 5’-дийодтиронин) – 0%; 3, 5, 3’, 5’-тетрайодтироуксусная кислота (тетрак) – 10-50%; 3, 5, 3’-трийодуксусная кислота (триак) – 25-35%; 3’-монойодтиронин, 3-монойодтиронин и, наконец, нейодированные тиронины. Метаболизм Т4 осуществляется не только путем последовательного дейодирования, но и разрыва связей между двумя ароматическими кольцами с образованием ДИТ. Около 15-20% тироксина инактивируется глюкуроновой и серной кислотами в печени с образованием глюкоронатов и сульфонатов, которые экскретируются с желчью. Лишь незначительная часть Т4 метаболизируется процессами дейодирования и сульфатирования в почках с последующей экскрецией метаболитов с мочой.
Хорошо известно, что тироидные гормоны необходимы для нормального роста и развития организма. Они контролируют образование тепла, скорость поглощения кислорода, участвуют в поддержании нормальной функции дыхательного центра, оказывают инотропный и хронотропный эффекты на сердце, увеличивают количество b-адренергических рецепторов в сердечной и скелетной мыщцах, жировой ткани и лимфоцитах, увеличивают образование эритропоэтина и повышают эритропоэз, стимулируют моторику желудочно-кишечного тракта, стимулируют синтез многих структурных белков в организме. У человека снижение биосинтеза и секреции этих гормонов приводит к задержке физического и психического развития, а также к нарушению дифференцировки скелета и ЦНС.
Содержание тироглобулина в сыворотке крови практически здоровых лиц составляет 5,1±0,5 нг/мл (J. Torrens и H. Burch, 1996) и повышается при злокачественных и некоторых других заболеваниях щитовидной железы. Показано, что в течение суток щитовидная железа секретирует 100 нмоль Т4, 5 нмоль Т3 и почти такое же количество обТ3. Около 80 нмоль Т4 конвертируется на периферии в Т3 (35 нмоль) или в обТ3 (45 нмоль). Оставшаяся часть Т4 (около 20 нмоль) инактивируется в основном в печени и небольшое количество в почках и экскретируется в виде тетрак и других соединений.
Концентрация Т4 в сыворотке крови составляет 8 мкг/100 мл (100 нмоль/л), период полураспада Т4 в плазме составляет около 7 дней; Т3 – 120 нг/100мл (1,8 нмоль/л), период полураспада в плазме около 1 дня и обТ3-около 40 нг/100мл (0,61 нмоль/л), период полураспада около 5 часов.
Биологическое действие гормонов щитовидной железы осуществляется путем регуляции белкового синтеза. Первым этапом в механизме действия тироидных гормонов является связывание гормона с соответствующим рецептором. Уже через 30 мин после внутримышечного введения Т3 связывается ядрами клеток печени и почек крыс. В дальнейшем происходит диссоциация Т3 из связи с ядерными рецепторами. Ядерный рецептор к Т3 клонирован. Различают три вида рецепторов к тироидным гормонам: a-1, a-2 и b-1 рецептор. a-1 рецептор имеет мол. м 47 кДа и состоит из 410 аминокислот. Ген, кодирующий этот рецептор, локализуется на 17-й хромосоме. a-2 рецетор тироидных гормонов практически не обладает способностью к комплексированию с тироидными гормонами и при определенных состояниях угнетает активность Т3. b-1 рецептор к тироидным гормонам с мол. м. 52 кДа включает 456 аминокислотных остатков и кодируется геном, локализованным на 3-й хромосоме. Каждый рецептор содержит 3 специфических домена: амино-терминальный, центральный ДНК-связывающий домен с двумя цистеин-цинковыми пальцами и карбокси-терминальный домен. Последний отвечает за комплексирование с гормоном. Амино-терминальный домен усиливает аффинность рецептора к гормону. После взаимодействия Т3 с рецептором образуется гормоно-рецепторный комплекс, который затем связывается со специальным элементом, чувствительным к тироидным гормонам, что приводит к повышению (или снижению) активности полимеразы II на Т3-чувствительном гене, что инициирует транскрипцию генов, отвечающих за образование мРНК и синтез соответствующих белков. Связывание Т3 ядерными рецепторами осуществляется без участия каких-либо цитоплазматических переносчиков, которые необходимы в механизме действия стероидных гормонов. Белки цитоплазмы могут связывать трийодтиронин, но их аффинность к Т3 значительно ниже по сравнению с таковой ядерных рецепторов.
Считается, что Т3 достигает ядра после серии неспецифических взаимодействий со связывающими белками цитоплазмы и проникает в ядро в “свободной”, а не в “связанной” форме. Функция белков цитозоля, таким образом, аналогична функции белков плазмы, связывающих тироидные гормоны. Ядерные рецепторы могут связывать не только Т3, но и Т4. Их способность связывать Т4 составляет лишь 1/10 или даже меньше от связывания Т3. Моно- и дийодтирозин не могут вытеснять (замещать) меченый Т3 из его с
|
 Главная
Главная